STRUCTURE OF P11 AND OF ITS COMPLEX WITH THE ANNEXIN II N-TERMINUS
S. Réty1,
J. Sopkova1, M. Renouard1,
S. Tabaries2, D. Osterloh3,
V. Gerke3, F. Russo-Marie2
& A. Lewit-Bentley1
1LURE ,
Centre Universitaire Paris-Sud, 91405 Orsay CEDEX, France,
2INSERM U332, I. C. G. M., 22 rue
Méchain, 75015 Paris, France
3Institute for Medical Biochemistry, ZMBE, University
of Münster, von-Esmarch-Straße 56, 48149 Münster, Germany,
Keywords: Calcium-binding protein; protein-peptide
complex.
Introduction: The S100 proteins, first identified in neuronal tissue, are small proteins of an average molecular weight of 10kd, formed by two EF-hand calcium binding motifs. The exact role of S100 proteins is not known but they may act in the regulation of cytoskeleton assembly and some are involved in brain pathologies (1). S100 proteins can form homo- or hetero-dimers with other S100 proteins in solution and some are covalently disulphide linked under physiological conditions, such as the secreted S100B dimer (2). The p11 protein, also known as calpactin I light chain or S100A10, is a distinct member of the S100 family since its two EF-hands are mutated in such a way it cannot bind any metal (3). Several S100 proteins have been shown to bind to members of the annexin family, in particular p11 to annexin II (4), calcyclin to annexin XI (5) and S100C to annexin I (6). Only in the latter two cases is the complex formation calcium dependent. Annexins are proteins which can bind negatively charged phospholipids in a calcium dependent manner and whose proposed functions include the regulation of membrane traffic, membrane-cytoskeleton contacts or ion channel (7). Some of their properties can be modulated by S100 protein interaction.
Results and Discussion: We have solved the crystal structure of p11 by multiple isomorphous replacement (Table 1). It shows a pair of helix-loop-helix EF-hand motifs linked by a flexible loop, with two molecules in the asymmetric unit forming a non-covalent dimer. This dimer is much tighter than the dimers described so far for S100 proteins, suggesting a central role for p11 in the formation of a hetero-tetramer with annexin II. p11 is the only member of the S100 family where mutations in the EF-hand motifs make calcium binding impossible. Our data indicate that the p11 structure resembles that of a S100 protein locked in the calcium-bound conformation. In spite of a stronger sequence homology with S100B, the p11 structure is closer to that of calbindin D9k, a protein thought to act as a calcium buffer and not in signalling. We found a disulphide-linked tetramer in the crystals, a form we find also in solution, showing it is not an artefact of crystallisation.
p11 was complexed with a tetra-decapeptide that corresponds to
the N-terminal sequence of annexin II and the crystal structure
of this complex was solved by molecular replacement (Table 1).
This is the first known structure of a complex of an S100 protein
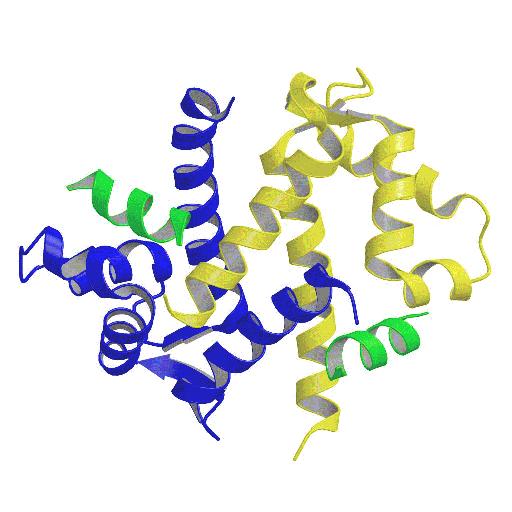
with its target peptide (Fig. 1). The peptide nestles in the
hydrophobic groove formed by the dimerisation of p11, in contact
with helices I and IV of both monomers and with loop L2 which is
stabilised in an a-helical conformation
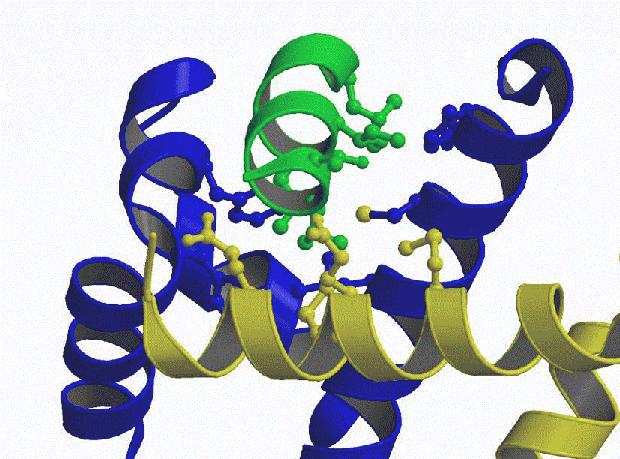
(Fig 2.). All the annexin II residues predicted from mutagenesis
studies are involved in contacts with the p11 dimer. The
N-terminus of the peptide lies in a cavity that cannot accomodate
any extension. The N-acetylation is important for the
stabilisation of the a-helix of the
peptide. The complex again forms a disulphide-linked dimer,
suggesting that the oxidation of p11 could have a physiological
relevance, as well as representing an important model for other
proteins of the S100 family, including those occuring
extracellularly in a disulphide-bridged form.
Table 1: Crystallographic parameters:
S.G. |
a(Å) |
b(Å) |
c(Å) |
ß(°) |
resol. |
Rsym |
R |
Rfree |
|
|---|---|---|---|---|---|---|---|---|---|
p11 |
I222 |
57.7 |
80.7 |
95.5 |
2.25 |
0.047 |
0.227 |
0.303 |
|
complex |
C2 |
80.6 |
56.4 |
64.3 |
114.5 |
2.45 |
0.060 |
0.32 |
0.4 |
|
|
| Figure 1. Schematic backbone tracing of the dimer of p11 (blue and yellow) with the bound peptide (green). Figure prepared with MOLSCRIPT (8). | Figure 2. Detail of the p11-peptide
interactions. The hydrophobic residues involved in the
interface are shown as balls. |